Anatomy & Physiology
Development of Polycarpa papillata after fertilisation is typical of ascidians, cleavage is holoblastic and bilateral which is a characteristic of deutrostomes, from this point in development chordate characteristics start to become evident (Ruppert et.al 2004). At gastrulation a large archenteron is formed at the posterior end and elongates, the mid-dorsal section of this elongate archenteron forms the notochord in later development (Ruppert et.al 2004). The neural tube is derived from the ectoderm which first differentiates into neural plates before curling over to form the neural tube (Ruppert et.al 2004). This development results in the formation ofa lecithotrophic larvae which lives in the water column for a maximum of thirty-six hours before it must settle and metamorphose, which limits the dispersal capabilities (Ruppert et.al 2004). During metamorphosis the larvae lose all of the evident chordate characteristics with the only common feature maintained in the adult being the gut morphology (Ruppert et.al 2004).
As filter feeders the ability to generate water current is a major part of their survival. In all tunicates this is accomplished by the use of lateral cilia that are located on the margins of the gill slits (Ruppert et.al 2004). The cilia are beat to generate this current and under normal conditions a body full of water is filtered per second which maintains the respiratory and metabolic requirements (Ruppert et.al 2004). To collect the food particles as they pass through the pharyngeal basket with the water a mucose net is produced which is transported to the esophagus and into the stomach (Ruppert et.al 2004). The reproductive system is attached to the latter end of the digestive tract surrounding the excurrent siphon (Kott, 1985). Respiration is also linked to the digestive tract with gas exchange occurring in the pharyngeal basket (Ruppert et.al 2004). See Figures 6 to 9.
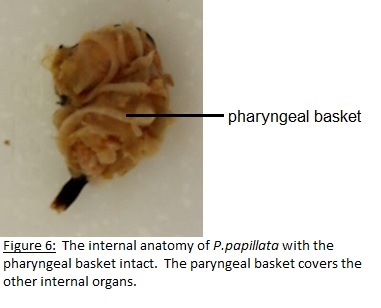
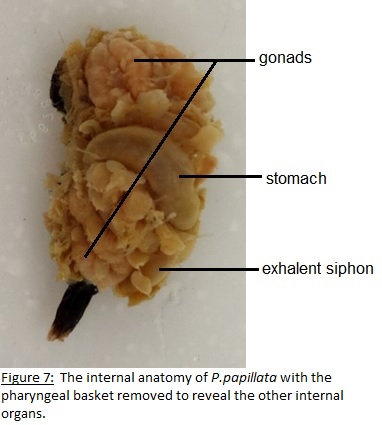
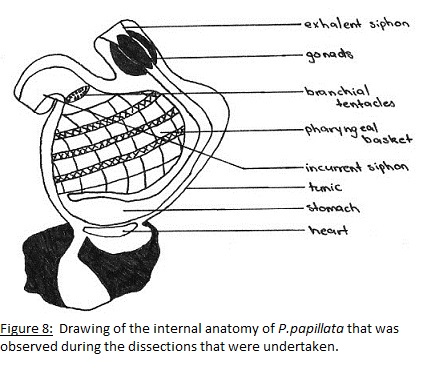
To investigate the behaviours that were observed when large particles entered the incurrent siphon (see Behaviour) dissections were undertaken. The aim was to discover what anatomical feature allows for the sense and rejection of large particles. Of the three specimens that were available two were dissected with the other having died in the aquarium before the dissection date. To dissect the individuals a cut was made from incurrent to excurrent siphon severing the nervous system and instantly killing the specimen. From there the tunic was cut from both siphons to attachment site and peeled back to reveal the internal anatomy (Fig. 6 and 7). It was thought that whatever feature was responsible for the rejection of particles would be located near the incurrent siphon so the incurrent siphon was removed and looked at under a dissecting microscope. It was found that the incurrent siphon was lined with tightly clustered tentacles(Fig. 9).

These are the branchial tentacles; these prevent large particles from entering the pharyngeal basket and eject them from the incurrent siphon which is the flicking behaviour (MacGinitie, 1939). It is when the large particles penetrate past this line of tentacles that the coughing behaviour is observed. The couching behaviour is the forcible ejection of the large particle from the pharyngeal basket by the body musculature (MacGinitie, 1939).
The tunic of P.papillata is extremely thick and rubber like due to the concentration of tunicin (Ruppert et.al 2004). This protects the individual and gives it support against water pressure, it is not moulted but grows with them throughout life (Shenkar and Swalla, 2011).
|